Sollys på puslespillet med prebiotisk RNA?
Carl Linné (psevdonym), January 24, 2024. Oversatt herfra
 I to tidligere artikler for Evolution News (her og her) har jeg argumentert mot sannsynligheten for å generere biologisk relevante polymerer (proteiner, RNA og DNA) i en prebiotisk verden. Mye av dette argumentet var basert på forespørselen om homokiralitet ved byggesteinene og de nøyaktige kjemiske bindingene nødvendig, for å oppnå nukleotider for RNA og DNA. Riktignok har noen forskere foreslått ordninger som er mer velsmakende for skeptikere av Opprinnelsen Til Liv (OTL) forskning. Derfor vil jeg her forklare de positive trinnene som nylig har blitt gjort for å løse gåten med hvordan RNA og DNA kan tenkes å oppstå i en abiotisk verden.
I to tidligere artikler for Evolution News (her og her) har jeg argumentert mot sannsynligheten for å generere biologisk relevante polymerer (proteiner, RNA og DNA) i en prebiotisk verden. Mye av dette argumentet var basert på forespørselen om homokiralitet ved byggesteinene og de nøyaktige kjemiske bindingene nødvendig, for å oppnå nukleotider for RNA og DNA. Riktignok har noen forskere foreslått ordninger som er mer velsmakende for skeptikere av Opprinnelsen Til Liv (OTL) forskning. Derfor vil jeg her forklare de positive trinnene som nylig har blitt gjort for å løse gåten med hvordan RNA og DNA kan tenkes å oppstå i en abiotisk verden.
I min tidligere artikkel -lenke , diskuterte jeg oppdagelsen av uracil som ble utvunnet fra prøver hentet fra en asteroide. Jeg gjorde det klart at dannelse av RNA (eller DNA) byggesteiner gjennom å koble ringstrukturerte nukleobaser (uracil, tymin, cytosin, adenin, guanin) til den ringbaserte sukker-riboser (eller deoksyribose for DNA) er et håpløst abiotisk foretak. Kjemien som trengs for å fullføre dannelse av de nødvendige byggesteinene i høyt utbytte, uten bruk av energiladede underlag og enzymer (som levende system bruker) er uholdbar. Flere OTL -forskere har anerkjent denne ulempen. De har valgt en annen vei, vurdert nedenfor, for å foreslå hvor biologisk relevante nukleotider kan ha blitt kjemisk produsert, ved bruk av bare startforbindelser som antas å være prebiotisk til stede.
Bilde 1. Sollys kan påvirke RNA-dannelse
Å takle syntese av pyrimidin -nukleosider (cytidin og uridin)
Det kjemiske skjemaet som er planlagt, benytter seg av en mekanisme for å bygge pyrimidinring-strukturer ab initio på ribose, i stedet for å prøve å koble dem til dette sukkeret. Hele den syntetiske prosessen er som følger:
-Ulike reaksjonsbetingelser i flertrinns-syntesene (forholdene er i fet tekst):
Trinn 1) Ribose + cyanamid / 0,2 m natriumkarbonat (pH ~ 10), 55grader C -> RAO
Trinn 2) Rao + cyanoacetylen / hydrosulfid i vasket formamid, UV -lys -> Midlertidig resultat A
Trinn 3) Intermedats A / fosfat, Anhydrous formamid, varme -> Alpha & Beta ribopyrimidines (pyrimidin ribonukleosider fullført)
Trinn 4) Intermedats A + 8 -Mercaptoadenine / tørr tilstand i magnesiumklorid, 150 grader C -> Mellomresultat B
Trinn 5) Intermedats B / bisulfat i fosfat (pH 7), 60 grader C eller sulfitt i fosfat (pH 8-10), varme, UV -lys -> mellomresultat C
Trinn 6) Mellomresultat C/ nitrolsyre (pH 4) -> Adenin og inosinprodukter (Purine Ribo og Deoxyribonucleosides fullført)
 Det første trinnet bruker cyanamid, en forbindelse som sannsynligvis er til stede på den tidlige jorden, for å reagere med ribose i en totrinns prosess som feller 5-karbon-sukkeret i den nødvendige furanosesring-konformasjonen.(1) Denne antatte RNA-forløperen, kalt RiboxAzOline (RAO) , har nå de tre første atomene som er bestemt til å danne den pyrimidin-ringen som allerede er festet til passende ribose karbon. Det er verdt å merke seg at ribose, som konkurrerer mot de tre andre mulige 5-karbon sukker, er det foretrukne underlaget i denne reaksjonen. Fra dette punktet fortsetter syntesen (2) via trinn 2 i tabellen, ved bruk av et annet enkelt abiotisk prospekt, cyanoacetylen, noe som resulterer i et mellomprodukt som lett omdannes til ribocytidin. I bare tre trinn ved bruk av enkle reaktanter er det første nukleosid nesten fullstendig i ganske høyt utbytte. Dette produktet må imidlertid være i beta -konfigurasjonen. Men dette styres enkelt ved eksponering for UV -lys med en konverteringsfrekvens så høyt som 74 prosent. UV -lys letter også oksidasjon av cytosin til uracil (ved mye lavere utbytte) som fullfører oppskriften for produksjon av begge pyrimidinnukleotider. Forfatterne i publikasjonsbyen ovenfor beskriver en alternativ rute til uridin, og tilbyr noen fordeler i utbytte ved å utføre reaksjonen med cyanoacetylen i nærvær av hydrosulfid.
Det første trinnet bruker cyanamid, en forbindelse som sannsynligvis er til stede på den tidlige jorden, for å reagere med ribose i en totrinns prosess som feller 5-karbon-sukkeret i den nødvendige furanosesring-konformasjonen.(1) Denne antatte RNA-forløperen, kalt RiboxAzOline (RAO) , har nå de tre første atomene som er bestemt til å danne den pyrimidin-ringen som allerede er festet til passende ribose karbon. Det er verdt å merke seg at ribose, som konkurrerer mot de tre andre mulige 5-karbon sukker, er det foretrukne underlaget i denne reaksjonen. Fra dette punktet fortsetter syntesen (2) via trinn 2 i tabellen, ved bruk av et annet enkelt abiotisk prospekt, cyanoacetylen, noe som resulterer i et mellomprodukt som lett omdannes til ribocytidin. I bare tre trinn ved bruk av enkle reaktanter er det første nukleosid nesten fullstendig i ganske høyt utbytte. Dette produktet må imidlertid være i beta -konfigurasjonen. Men dette styres enkelt ved eksponering for UV -lys med en konverteringsfrekvens så høyt som 74 prosent. UV -lys letter også oksidasjon av cytosin til uracil (ved mye lavere utbytte) som fullfører oppskriften for produksjon av begge pyrimidinnukleotider. Forfatterne i publikasjonsbyen ovenfor beskriver en alternativ rute til uridin, og tilbyr noen fordeler i utbytte ved å utføre reaksjonen med cyanoacetylen i nærvær av hydrosulfid.
Bilde 2. Vurdering omkring livets opprinnelse (O. Grasso)
Å takle syntese av purin [deoksy] nukleosider ([deoxy] adenosin og [deoxy] inosin)
For å takle produksjonen av purinnukleotider, reagerte en annen formodet kjemisk forløper mulig i en abiotisk setting (trinn 4) Ved UV -bestråling der den biologisk relevante isomeren gir ut gir nærmere 50 prosent. Behandling av DA med nitgallsyre blir gitt delvis konvertering til beta-DeoksyInosin (DI). Etterfølgende forskning fra denne gruppen demonstrerte muligheten for å produsere de tilsvarende purin ribonukleosider (A og i) ved å utføre den første reaksjonen i alkalisk natriumsulfitt i stedet for bisulfat.(4) Utbyttet av de ønskede produktene er betydelig lavere enn de som ble funnet for pyrimidin (( 10-40 prosent mot 40-50 prosent). Oppsummert har disse kjemiske syntetiske studiene under hypotetiske prebiotiske forhold vist respektable utbytter for beta -versjonene av ribo -pyrimidiner og puriner, så vel som for beta -versjoner av Deoxyribo Puriner, som representerer de kanoniske byggesteinene som ble brukt i biologi.
Homokiralitets-utfordringen
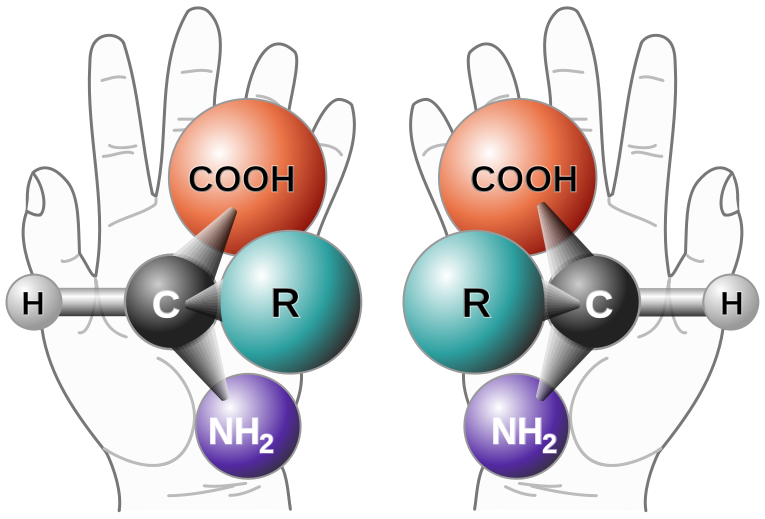 Ingen av studiene som er angitt ovenfor, skiller mellom de to stereoisomerer av ribose. Visstnok burde L-ribose ha vært like til stede med D-ribose i en prebiotisk verden, så RAO-forløpere ville på samme måte være en racemisk blanding. Et annet laboratorium undersøkte en innovativ tilnærming som utnyttet det faktum at kirale molekyler kan samkjøre seg med et magnetfelt på grunn av spinntilstander til alle dets elektroner.(5) Enantiomere par av en forbindelse vil utvise spinntilstand-tilpasning, med motstridende magnetisk polaritet. Dette prinsippet er kjent som kiral-indusert spinnselektivitet (CISS). Fordi magnetitt forventes å være rikelig i sedimentet av vannmasser på den primordiale jorden, ble det antatt at jordens magnetfelt ville tjene til å magnetisere det jernholdige sedimentet, og ga et middel til å fortrinnsvis absorbere RAO-enantiomerer i ulike Ciss. Referanseeksperimentene ble utført i petriskåler som simulerte denne designen, og plasserte en magnet under den uoppløselige magnetitten. Den første utvelgelsesrunden førte til slutt til RAO-krystallisering på magnetittoverflaten, og demonstrerte en betydelig berikelse i enten D-eller L-RAO avhengig av magnetisk polaritet.
Ingen av studiene som er angitt ovenfor, skiller mellom de to stereoisomerer av ribose. Visstnok burde L-ribose ha vært like til stede med D-ribose i en prebiotisk verden, så RAO-forløpere ville på samme måte være en racemisk blanding. Et annet laboratorium undersøkte en innovativ tilnærming som utnyttet det faktum at kirale molekyler kan samkjøre seg med et magnetfelt på grunn av spinntilstander til alle dets elektroner.(5) Enantiomere par av en forbindelse vil utvise spinntilstand-tilpasning, med motstridende magnetisk polaritet. Dette prinsippet er kjent som kiral-indusert spinnselektivitet (CISS). Fordi magnetitt forventes å være rikelig i sedimentet av vannmasser på den primordiale jorden, ble det antatt at jordens magnetfelt ville tjene til å magnetisere det jernholdige sedimentet, og ga et middel til å fortrinnsvis absorbere RAO-enantiomerer i ulike Ciss. Referanseeksperimentene ble utført i petriskåler som simulerte denne designen, og plasserte en magnet under den uoppløselige magnetitten. Den første utvelgelsesrunden førte til slutt til RAO-krystallisering på magnetittoverflaten, og demonstrerte en betydelig berikelse i enten D-eller L-RAO avhengig av magnetisk polaritet.
Etter å ha samlet krystallene, redissolvert dem i buffer og gjentatt det magnetittinduserte utvalget, resulterte den andre runden med krystallisering i optisk ren RAO. Fra eksperimenter som disse ble det foreslått at homokiralitet av RAO kunne vært oppnådd på den prebiotiske jorden der en liten enantiomer-ubalanse induseres i soner for å beholde fortrinnsvis en av de to RAO -serreoisomerer. Med dette som forekom nær kysten, er vannstrømmen til RAO -krystaller, etterfulgt av en påfølgende tørrfase når strandlinjen går tilbake. Når vannet stiger til dette nivået igjen, omkrystalliserer RAO seg til et større enantiomert overskudd. Det er sinus magnetitten hadde blitt betinget forbedret magnetisk i forrige runde gjennom CISS-effekten.(6) Det ble antydet at denne gjentatte syklusen av adsorpsjon, krystallisering og spin-oppstilling på magnetitt overflaten, kan tjene som en positiv feedback-loop, som fører til den ultimate kostnaden for krystalliserende homokiralt RAO.
Bilde 3. Homokiralitet demonstrert (speilvendte molekyler)
Mulighet for denne kjemien ble demonstrert, men er det sannsynlig under prebiotiske forhold?
For først å betrakte forslaget om å oppnå homokiralitet av RAO, er det stilt svake forutsetninger for å akseptere denne prestasjonen. Forfatterne bemerket at tidspunktet for krystallisering er viktig. Det skyldes at hvis krystaller får lov til å vokse i for lang tid, reduseres enantiomene overskudd og dermed reduseres den kirale spinn-tilstandstreningen av magnetitt, eller ødelegges. Det vil ta en god del flaks for å ha den syklusen spontant gjentatt til riktig tid. Det kan også forventes at differensialsoner langs kysten, ville velge hver sin enantiomer av RAO.(6) Til tross for individuelle homokirale krystaller over større område, er de motsatte stereoisomerer også krystallisert. Så hvordan forblir de atskilt før og etter å ha gjennomgått påfølgende reaksjoner, noe som fører til nukleotidsyntese? Dette kan lett oppnås i laboratoriet, og der holde krystaller og deres nedstrømningsprodukter isolert. Men det er ingen garanti for at de vil forbli isolert under abiotisk situasjon.
 Når det gjelder syntese av nukleosider fra RAO, spiller flere forhold inn: 1) Multitrinns syntese reduserer utbyttet av sluttprodukter og øker sannsynlighetene for å generere biprodukter som kan konkurrere om inkorporering i RNA-byggesteiner. Spesielt utbyttet av purin -nukleotider lider av kravet om ytterligere trinn som trengs i syntesen. 2) De fleste reaksjoner krever dispensatoriske forhold. Hvordan blir reaktantene i et abiotisk miljø sekvensielt inkorporert under disse forskjellige reaksjonsbetingelsene (trinn 1-6)? I laboratoriet er dette selvforklarende, men det er ikke en triviell sak når kjemi blir overlatt til stokastiske hendelser. 3) Spådommer om det kjemiske repertoaret til en prebiotisk jord er stort sett underbygget. Støtte for slike spådommer kan potensielt komme fra å analysere prøver fra andre utenomjordiske organer som det ble gjort med asteroiden Ryugu. Henting av prøver fra Mars ville være et spesielt spennende sted å utforske. Uten denne typen bekreftende data kan våre antagelser om prebiotisk kjemi ganske enkelt famle i blinde. 4) UV -lysets rolle er sannsynligvis kritisk for dannelsen av disse RNA -byggesteinene. Dette kan være et tveegget sverd da UV-lys også kan skade puriner og pyrimidiner på flere måter. 5) Et prebiotisk syntetese skjema som fører til guanin er fremdeles ikke utdypet. I stedet foreslår de at inosin, som er inkludert i ordningene som er presentert her, kan erstatte guanin sinus, det er i stand til å basere par med cytosin (som guanin gjør). Inosin er imidlertid mer beslektet med å være en joker, da det også danner baserepar med uracil og tymin. Slik promiskuøs baseparing vil upålitelig begrense den funksjonelle spekteret av RNA/DNA -polymer og endre den strukturelle dynamikken som vi nå vet er viktig for de biologiske rollene til mange RNA -er.
Når det gjelder syntese av nukleosider fra RAO, spiller flere forhold inn: 1) Multitrinns syntese reduserer utbyttet av sluttprodukter og øker sannsynlighetene for å generere biprodukter som kan konkurrere om inkorporering i RNA-byggesteiner. Spesielt utbyttet av purin -nukleotider lider av kravet om ytterligere trinn som trengs i syntesen. 2) De fleste reaksjoner krever dispensatoriske forhold. Hvordan blir reaktantene i et abiotisk miljø sekvensielt inkorporert under disse forskjellige reaksjonsbetingelsene (trinn 1-6)? I laboratoriet er dette selvforklarende, men det er ikke en triviell sak når kjemi blir overlatt til stokastiske hendelser. 3) Spådommer om det kjemiske repertoaret til en prebiotisk jord er stort sett underbygget. Støtte for slike spådommer kan potensielt komme fra å analysere prøver fra andre utenomjordiske organer som det ble gjort med asteroiden Ryugu. Henting av prøver fra Mars ville være et spesielt spennende sted å utforske. Uten denne typen bekreftende data kan våre antagelser om prebiotisk kjemi ganske enkelt famle i blinde. 4) UV -lysets rolle er sannsynligvis kritisk for dannelsen av disse RNA -byggesteinene. Dette kan være et tveegget sverd da UV-lys også kan skade puriner og pyrimidiner på flere måter. 5) Et prebiotisk syntetese skjema som fører til guanin er fremdeles ikke utdypet. I stedet foreslår de at inosin, som er inkludert i ordningene som er presentert her, kan erstatte guanin sinus, det er i stand til å basere par med cytosin (som guanin gjør). Inosin er imidlertid mer beslektet med å være en joker, da det også danner baserepar med uracil og tymin. Slik promiskuøs baseparing vil upålitelig begrense den funksjonelle spekteret av RNA/DNA -polymer og endre den strukturelle dynamikken som vi nå vet er viktig for de biologiske rollene til mange RNA -er.
Bilde 4. James Tour er klar angående OTL
Virkelig en ekstraordinær løsning
 Jeg (Linné) etter pris på kvaliteten på arbeidet som gikk i å utvikle kjemiske ordninger for å syntetisere purin- og pyrimidin -nukleotidbaser, og oppnå den obligatoriske stereo- og kjemiske spesifisiteten til furanosyl ribose etablert i levende systemer. Dette er virkelig en ekstraordinær løsning ved bruk av påståtte prebiotiske forhold. Etter min mening har andre forsøk på å møte denne utfordringen ikke kommet i nærheten av å oppnå et slikt nøyaktighetsnivå. Dessverre er det viktigste problemet hvordan dette kan fullføres uten en guidet mekanisme for å styre hver påfølgende reaksjon og opprettholde respektable utbytter, langs hele prosessen.
Jeg (Linné) etter pris på kvaliteten på arbeidet som gikk i å utvikle kjemiske ordninger for å syntetisere purin- og pyrimidin -nukleotidbaser, og oppnå den obligatoriske stereo- og kjemiske spesifisiteten til furanosyl ribose etablert i levende systemer. Dette er virkelig en ekstraordinær løsning ved bruk av påståtte prebiotiske forhold. Etter min mening har andre forsøk på å møte denne utfordringen ikke kommet i nærheten av å oppnå et slikt nøyaktighetsnivå. Dessverre er det viktigste problemet hvordan dette kan fullføres uten en guidet mekanisme for å styre hver påfølgende reaksjon og opprettholde respektable utbytter, langs hele prosessen.
Utover den innledende syntese av byggesteiner for nukleinsyre, er det mer signifikante bekymringer når nukleotidkjeder er samlet. Tilstøtende pyrimidiner er veldig utsatt for å danne flere typer kjemiske lesjoner, fremkalt av UV -lys. Uten en serie enzymer for å reparere slike lesjoner, er virkningene for det skadede RNA/DNA -molekylet dårlig. I nåværende biologiske systemer resulterer de i mutasjoner på grunn av replikasjonsfeil. I en prebiotisk verden ville reparasjons-enzymer selvfølgelig ikke eksistere. Disse avsluttende merknadene kaster lys (unnskyld ordspillet) over fremtidige hensyn til RNA/DNA -polymer i en prebiotisk verden. Hvis man lurer på hvorfor OTL-vitenskap har sine skeptikere, avslører upretensiøse komplikasjoner som de her, tvil som oppstår når vi tenker på (vitenskaps) feltet.
For Referanser se nederst i originalen-lenke.
Bilde 5. Hvordan arrangere byggestenene i cellen?
Carl Linné
Carl Linné (et pseudonym) fikk sin doktorgrad i biokjemi og ledet et akademisk laboratorium som undersøkte strukturen/funksjonsforholdene til medikamentreseptorer.
Oversettelse, via google oversetter, og bilder ved Asbjørn E. Lund